Микрогаметогенез у межвидового триплоидного гибрида вишни Cerasus vulgaris Mill x Padellus maxi-moviczii Erem. et. Jushev.
УДК 576.354.4:575.127.2:634.23 2007
Авторы: Седышева Г. А., Джигадло Е. Н., Горбачева Н. Г.
В связи с широким распространением в последние десятилетия таких опасных болезней как коккомикоз и монилиоз, в ряде зон вишня утратила свое промышленное значение. В этой связи, привлечение в селекцию вида вишня Максимовича или, по данным современной систематизации, Padellus maximoviczii Erem et Jushev (Юшев, 1993), которая обладает иммунитетом к обеим этим болезням, имеет большое значение для восстановления статуса этой культуры. Гибриды, полученные от скрещивания с этим видом, практически не поражаются ни коккомикозом, ни монилиозом. Примером может служить отдаленный межвидовой гибрид вишни № 85023 Cerasus vulgarus Mill. x Padellus maximoviczii Erem. еt Jushev,полученный в лаборатории селекции косточковых культур ВНИИСПК Е. Н. Джигадло. В 2000 году этот гибрид был выделен как донор устойчивости к монилиозу и коккомикозу.
Гибрид №85023 характеризуется среднепоздним сроком цветения и созревания. Средняя степень цветения 3,5 балла. Степень плодоношения 10-летнего дерева – 1 балл (рис. 1).
Рис. 1. Плодоношение гибрида вишни 85023 (C. vulgaris x P. maximoviczii).

Плоды мелкие – 2,5 г, почти черные, сок темно-красный. Внешний вид – 3,8 балла. Вкус кисло-сладкий, 3,5 балла. Степень подмерзания за все годы изучения 0 баллов, за исключением зимы 2006 года, когда было отмечено подмерзание на 1 балл. Поражение коккомикозом – 0 баллов, монилиозом – 0 баллов за все годы наблюдения.
Слабое плодоношение гибрида 85023 объясняется тем, что этот гибрид имеет триплоидный набор хромосом 2n=3х=24. Как известно, триплоидные гибриды вишни в большинстве случаев либо полностью стерильны, либо характеризуются слабой плодовитостью (Жуков, Харитонова, 1977; Машкина, 1977 и др.). Для производственных целей стерильные и даже слабоплодовитые формы ценности не представляют. Однако, вслучае необходимости использования в селекции, слабоплодовитые формы могут представлять несомненный интерес, если несут в себе такие положительные качества, как иммунитет к таким опасным заболеваниям как коккомикоз и монилиоз. Как раз этими качествами и обладает триплоидная гибридная форма вишни 85023. Поэтому изучение хода микрогаметогенеза и определение качества формируемых этой формой гамет позволит установить возможность использования ее в селекции для получения сортов вишни, устойчивых к данным заболеваниям. Для изучения хода мейоза цветковые почки фиксировали в уксусном алкоголе (3 : 1). Фиксацию проводили в условиях лаборатории со срезанных ветвей, поставленных в вегетационные сосуды с водой. Фиксация темпоральная, на протяжении нескольких дней с момента появления зеленого конуса и до распускания цветка. Из фиксированного материала готовили временные давленые препараты ацетогематоксилиновым методом (Топильевская, Лучникова, Чувашина, 1975). Препараты изучали на микроскопе Biolar, при увеличении 10 х 1,5 х 40, 10 х 1,5 х 100.
Результаты исследований
Анализируя состояние генеративной сферы у отдаленного триплоидного гибрида вишни 85023 (С. vulgaris Mill x Рadellus maximoviczii Erem. et Jushev) следует отметить, что нарушения в спорогенной ткани наблюдаются еще до начала мейоза. Так, на стадии профазы среди нормальных клеток вторичного археспория встречаются клетки, где вместо одного ядра, как это бывает в норме, находится по 2 ядра нормальной величины. Иногда число дополнительных ядер достигает 2, 3 идаже 5, но более мелких чем основные. Причем размер самой спорогенной клетки превышает обычный примерно в два раза. Как будет видно далее, последствия этого нарушения прослеживаются на всех последующих этапах микроспорогенеза.
Ход мейоза у данного триплоидного гибрида сопровождается многочисленными нарушениями на всех последовательных стадиях(табл. 1). Число этих нарушений варьирует от 43,3% на стадии тетрад до 80,2% на стадии анафаза – I.
1. Характеристика мейоза при микроспорогенезе у триплоидного гибрида вишни 85023 (C. Vulgarus x P. maximoviczii)
Стадии мейоза | Всего изучено клеток | В том числе | ± m | |||
нормальных | с нарушениями | |||||
шт. | % | шт. | % | |||
Метафаза – I | 247 | 121 | 49,0 | 126 | 51,0 | 10,1 |
Анафаза – I | 177 | 35 | 19,8 | 142 | 80,2 | 8,9 |
Телофаза - I | 214 | 78 | 36,4 | 136 | 63,6 | 10,8 |
Метафаза – II | 204 | 66 | 32,4 | 138 | 67,6 | 10,7 |
Анафаза – II | 186 | 93 | 50,0 | 93 | 50,0 | 13,4 |
Телофаза – II | 231 | 110 | 47,6 | 121 | 52,4 | 10,8 |
Тетрады | 416 | 236 | 56,7 | 180 | 43,3 | 5,9 |
Одноядерная пыльца | 518 | 312 | 60,2 | 206 | 39,8 | 4,6 |
Спектр нарушений довольно широк. На разных стадиях мейоза наблюдали от трех до восьми типов нарушений (табл. 2).
Следует отметить, что первое – гетеротипическое деление мейоза протекает с несколько большим числом нарушений, нежели второе - гомеотипическое. Однако, если в первом делении на каждой стадии наблюдается не более 5 типов нарушений, то во втором делении это разнообразие значительно больше. Например,во время анафазы – I число типов нарушений достигает восьми (табл. 2).
Наиболее распространенным типом нарушений на стадии метафаза – I является забегание одной или нескольких хромосом к одному или обоим полюсам веретена деления (86,5 % от общего числа нарушений). Многочисленные забегания отдельных хромосом в метафазе – I, по всей вероятности, можно объяснить тем, что бивалентная конъюгация (формула метафазы – I М – I = 12 II) отмечена только в 15,4 % клеток. В остальных случаях (84,6 %) наблюдается наличие 2-6 унивалентов. Общий характер конъюгации хромосом у формы 85023 представлен в таблице 3.
2. Типы нарушений на последовательных стадиях мейоза у триплоидного гибрида вишни C. vulgarus x P. maximoviczii
Стадия мейоза | Общее число нарушений | Типы нарушений | Кол-во клеток | % от общего числа наруш. |
Метафаза – I | 126 | забегание 1-4 хромосом | 109 | 86,5 |
выброс 1-3 хромосом в цитоплазму | 6 | 4,8 | ||
2-х ярусное распределение хромосом | 3 | 2,4 | ||
диффузное распределение хромосом | 1 | 0,8 | ||
Комбинация из 2-х типов | 7 | 5,6 | ||
Анафаза – I | 142 | отставание 1-7 хромосом | 131 | 92,3 |
комбинация из 2-х типов | 5 | 3,5 | ||
ценоциты | 2 | 1,4 | ||
неравные анафазные группы | 3 | 2,1 | ||
выброс | 1 | 0,7 | ||
Телофаза - I | 136 | микроядра (1-4) | 132 | 97,1 |
асинхронное формирование ядер | 1 | 0,7 | ||
одноядерный микроспороцит | 2 | 1,5 | ||
| МКП в 2 раза крупнее нормы, с 4-мя крупными ядрами | 1 | 0,7 | ||
Метафаза – II | 138 | забегания | 75 | 54,3 |
выбросы (1-4) | 33 | 23,9 | ||
неравные метафазные группы | 5 | 3,6 | ||
сверхчисленные веретена | 4 | 2,8 | ||
диффузное распределение хромосом | 2 | 1,4 | ||
комбинация из 2-х типов | 17 | 12,2 | ||
ценоцит | 2 | 1,4 | ||
Анафаза - II | 93 | отставания 1-4 хромосом | 62 | 66,7 |
| выбросы (1–3) | 10 | 10,8 | ||
комбинация из 2-х типов | 9 | 9,7 | ||
сверхчисленные веретена (3-4) | 5 | 5,4 | ||
мост | 1 | 1,1 | ||
неравное распределение хромосом | 2 | 2,1 | ||
диффузное распределение хромосом | 2 | 2,1 | ||
| ценоцит | 2 | 2,1 | ||
Телофаза -II | 121 | микроядра (1-4) | 108 | 89,3 |
2 или 3 ядра + микроядра | 4 | 3,3 | ||
ценоцит (от 6 до 20 ядер) | 9 | 7,4 | ||
| Тетрады | 180 | монада | 1 | 0,5 |
диада | 3 | 1,7 | ||
триада | 7 | 3,9 | ||
сверхчисленные микроспоры от 1 до 6 | 169 | 93,9 |
3. Характер конъюгации хромосом у триплоидного отдаленного гибрида вишни 85023
Показатели | Всего изучено МКП | Ассоциации | |||
Всего | в том числе | ||||
унивал. | бивален. | тривал. | |||
Изучено МКП (шт.) | 13 | ||||
Общее число ассоциаций | 175 | 43 | 127 | 5 | |
Число ассоциаций в среднем на 1 МКП | 13,5 | 3,3 | 9,8 | 0,4 | |
Варьирование | 12-15 | 0-6 | 8-12 | 1-2 | |
Процент | 100 | 24,6 | 72,6 | 2,8 | |
Таким образом, анализ данных таблицы 3 свидетельствует, что биваленты составляют основную часть ассоциаций хромосом в метафазе – I . В среднем на клетку приходится 9,8 бивалентов, унивалентов – 3,3 и тривалентов – 0,4. Ассоциаций валентностью выше трех у формы 85023 не наблюдали.
На стадии анафазы первого мейотического деления наиболее многочисленным типом нарушений является отставание 1-7 хромосом в процессе анафазного движения в центре ахроматинового веретена (92,3 %).
Телофаза – I характеризуется наличием сверхчисленных ядерили микроядер. Как правило, их насчитывается от 1 до 4. Такие микроспороциты составляют 97,1 % от общего числа нарушений.
Во втором мейотическом делении в метафазе – II преобладающим типом нарушений являются забегания и выбросы. В общей сложности они составляют 78,2 %. Для анафазы – II характерны, в основном, отставания в центре ахроматинового веретена, выбросы хромосом за пределы веретена, сверхчисленные фигуры деления. В общей сложности эти типы нарушений составляют более 80 % от общего числа нарушений.
На стадии телофазы – II самым многочисленным типом нарушений является наличие сверхчисленных ядер и микроядер в 89,3 % клеток. Следствием этого является формирование полиад вместо нормальных тетрад (рис. 2 п). Число микроспор в спораде колеблется от 5 (пентада) до 10 (декада) (рис. 2 с). Отмечены также случаи формирования спорад с числом микроспор меньше нормы: триад, диад (рис. 2 Р) и даже монад. Кроме отмеченных типов нарушений в гетеротипическом делении мейоза встречаются, но со значительно меньшей частотой, двухъярусное расположение хромосом в экваториальной плоскости, диффузное расположение хромосом по всему микроспороциту, одновременно несколько типов нарушений в одном микроспороците, асинхронное развитие ядер в телофазе – I (рис. 2 ж).
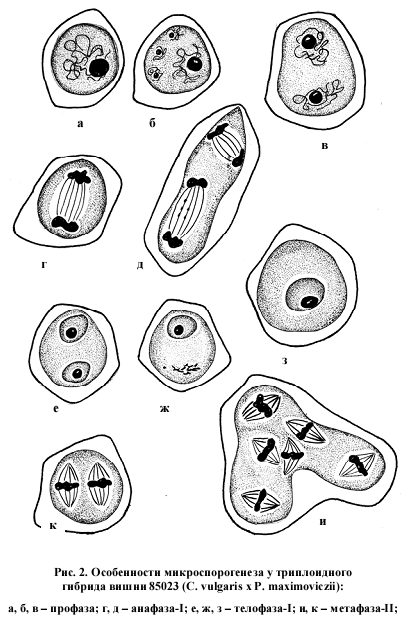
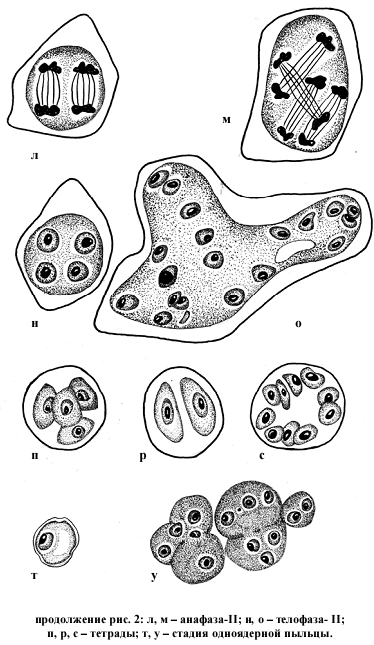
В гомеотипном делении также наблюдается диффузное распределение хромосом или по всему веретену, или по всему микроспороциту, комбинация из двух или более типов нарушений в одной клетке, сверхчисленные фигуры деления.
Некоторые нарушения подобного рода ранее наблюдали у отдаленных гибридов между вишней обыкновенной и вишней войлочной Н. П. Чувашина, В. Г. Касьянова (1967), Г. А. Седышева, А. Ф. Колесникова (1979). Однако внастоящем исследовании у отдаленного гибрида вишни 85023 (C. vulgarus x P. maximoviczii) при развитии спорогенных клеток отмечено нарушение, которое отличает эту форму от ранее изученных. Как уже упоминалось ранее, в спорогенной ткани еще до начала мейоза наблюдаются очень крупные материнские клетки пыльцы (МКП), которые в два раза крупнее обычных и содержат либо два нормальных ядра, либо одно нормальное ядро и 2 ядра меньшего размера (рис. 2 а, б, в). Иногда в нормальных МКП наряду с одним нормальным ядром присутствуют еще 2, 3 или 5 микроядрышек. Далее, почти на всех последующих стадиях отмечено наличие гигантских образований. В анафазе – I это крупный микроспороцит с двумя нормальными веретенами (рис. 2. г, д), в телофазе – I очень крупный одноядерный микроспороцит (рис. 2. з). На стадии метафаза – II очень крупные микроспороциты неправильной формы с двумя или тремя парами веретен нормальной конструкции и нормального размера (рис. 2. и, к). В анафазе – II это также гигантские образования со сверхчисленными веретенами (рис. 2. л, м). Особенно много подобных структур на стадии телофаза – II. Микроспороциты здесь представляют собой ценоцитные образования в большинстве случаев неправильной формы с числом ядер от двух (очень крупных) до 6, 7, 8, 9, 12 и даже 20 (рис. 2. н, о). Ядра в этих мультиядерных микроспороцитах обычного размера. В общей сложности число таких структур на стадии телофаза – II составляет 4,8 %. Позднее, на стадии тетрад наличие гигантских структур отмечается очень редко. Возможно, что перегруженность генетическим материалом отрицательно сказывается на слаженности метаболических процессов и содержимое таких ценоцитных образований в большинстве случаев постепенно подвергается деструкции. Мы наблюдали только один случай формирования микроспор в таком ценоцитном образовании. В пыльнике, рядом с одноядерной пыльцой находилась группа клеток, которые имели вид микроспор из только что распавшихся микроспороцитов. Но в данном случае они не были обособленными, а представляли собой как бы конгломерат слипшихся микроспор. Причем, эти микроспоры были разной величины и содержали разное число ядер (рис. 2. т, у).
На стадии молодой одноядерной пыльцы крупные пыльцевые зерна (в 2-4 раза крупнее обычных) составляют 11,9 %, мелкие (в два раза меньше обычных) – 27,9 %. Одномерная, среднего размера пыльца составляет 60,2 %. Но несмотря на это, количество жизнеспособной пыльцы будет значительно ниже, так как следует учитывать, что гаметы со сбалансированным числом хромосом составят значительно меньший процент. Подтверждением тому может служить анализ анафазы- II. Распределение хромосом по анафазным группам очень нарушено. Гаметы будут содержать самые разнообразные числа хромосом, в большинстве случаев анеуплоидные (табл. 4).
4. Характеристика анафазы - II у триплоидного отдаленного гибрида вишни 85023 (C. vulgarus x P. maximoviczii)
Формула анафазы - II | Всего гамет | В том числе: | |
с n = 8 шт./% | с n = 16 шт./% | ||
12 + 12 + 12 + 12 | 4 | - | - |
8 + 4 + 9 + 13 + 14 | 5 | 1 | - |
8 + 8 + 2 + 16 + 14 | 5 | 2 | 1 |
13 + 11+ 12 + 12 | 4 | - | - |
7 + 6 + 17 + 18 | 4 | - | - |
12 + 12 + 11 + 11 + 1 + 1 | 6 | - | - |
14 + 14 + 2 + 8 + 10 | 5 | 1 | - |
2 + 8 + 8 + 15 + 15 | 5 | 2 | - |
8 + 8 + 16 + 3 + 13 | 5 | 2 | 1 |
Всего: | 43 | 8/18,6 | 2/4,7 |
Таким образом, 76,7 % гамет будут содержать анеуплодные числа хромосом, а следовательно, такая пыльца, вероятнее всего, будет нежизнеспособна. Лишь 18,6 % гамет будут иметь гаплоидное число хромосом n = х = 8 и 4,7 % - диплоидное n = 2х = 16. Наличие таких сбалансированных гамет делает эту форму пригодной для использования в селекции с целью получения устойчивых к коккомикозу и монилиозу сортов вишни.
Подобное явление, когда наряду с образованием в подавляющем большинстве анеуплоидных гамет, регулярно формируется некоторое количество жизнеспособных, сбалансированных гамет, отмечено нами ранее у частично плодовитой формы вишни (C.vulgaris х C. sachalinensis) (Седышева, Джигадло, Князева, 1996).
Такая повторяемость этого явления позволяет сделать предположение, что оно закономерно и находится под генетическим контролем.